
Corals and many other reef invertebrates such as giants clams, gorgonians, sponges and hydroids live in a symbiotic relationship with dinoflagellates, also referred to as "zooxanthellae", that provide them with an important part of their daily energy requirement through photosynthesis. The genotype of dinoflagellates (Symbiodinium) plays an important role in determining the environmental tolerance range of their host. The genus Symbiodinium consists of nine broad genetic groups or clades that are genetically highly distinct (clades A-I).
Corals and many other reef invertebrates such as giants clams, gorgonians, sponges and hydroids live in a symbiotic relationship with dinoflagellates, also referred to as "zooxanthellae", that provide them with an important part of their daily energy requirement through photosynthesis. When the symbiosis breaks down, which may be caused by a range of factors but is most commonly due to an increase in sea surface temperature (SST), the dinoflagellate symbionts are lost and the host appears bleached. Bleaching reduces growth rate and when the bleached state of a colony persists it may lead to death.
The genotype of dinoflagellates (Symbiodinium) plays an important role in determining the environmental tolerance range of their host. The genus Symbiodinium consists of nine broad genetic groups or clades that are genetically highly distinct (clades A-I). So far, all clades except E, H and I have been found in corals although on the Great Barrier Reef most corals harbour Symbiodinium types from clade C and to a lesser extent clade D. Clade E is found in anemones and clade F, G, H and I are common among foraminifera. Each of these clades contains genetically and ecologically distinct subcladal types or strains. These types can, but do not necessarily have to be, different Symbiodinium species. Subcladal types often only differ by a few nucleotides. Yet, they may vary distinctively in their physiological characteristics; from differences in symbiont size and pigment composition to differences in heat stress tolerance.

Description
Bleached Fungia spp. next to other unbleached Fungia spp. solitary corals. The bleached coral may have had a clade of zooxanthellae alga that is more temperature sensitive than the unbleached coral.
The functional and ecological differences have been well described for a few Symbiodinium types, for instance subcladal type C1 is known to be more temperature tolerant than type C3 in their Acropora hosts resulting in differences in their bleaching susceptibility. However, the majority of the subcladal types have not been investigated in this way. Corals containing more temperature tolerant symbionts may not only resist the effects of environmental changes for longer periods of time but are also potentially more resilient and have increased survival rates after stress conditions subside. The abundance and distribution of stress tolerant Symbiodinium types will therefore provide useful information on how coral communities respond to climate change. Besides symbiont identity many other factors (for example the type of host, growth and reproduction) determine the physiological tolerance of a coral and its ability to cope with changing environmental conditions.
Despite the important role Symbiodinium might play we currently have limited knowledge of the distribution ranges of Symbiodinium on the Great Barrier Reef (GBR). In order to increase our knowledge a database of Symbiodinium diversity on the GBR compiled from published papers (see database references) was established as part of a large research program 1.1 entitled “Risk, Resilience and Response Atlas for the Great Barrier Reef: Vulnerability to Coral Bleaching Project Objective.” The goal of project 1.1 is to define the vulnerability of GBR corals to climate change and extend this knowledge along the geographical extent of the GBR. The database presents an overview of the Symbiodinium types that have thus far been found across the GBR and can be accessed through e-Atlas. Understanding Symbiodinium diversity and the geographical and environmental ranges of the different host/symbiont combinations will help us provide a better estimate of how vulnerable corals on the GBR are to bleaching.
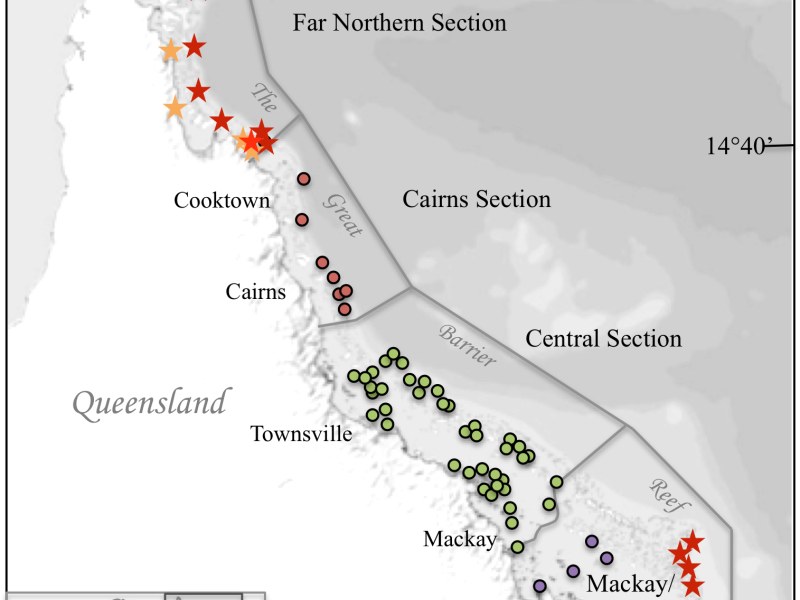
Figure 1: Map showing the sample locations on the Great Barrier Reef. Circles indicate sites extracted from published literature (different colours indicate different sections of the GBR). Stars indicate additional collection sites (inshore = orange, offshore = red).
View sites in Google Earth
Existing Symbiodinium collections are largely restricted to central and mid-shelf GBR locations (Figure 1, database collection sites are indicated by dots) and substantial variability in symbionts was highlighted by these studies. As such it is unlikely that these studies have captured the full diversity of symbionts present on the GBR. We therefore extend the existing collection by sampling a wide variety of Symbiodinium bearing hosts (Scleractinea, Alcyonacea, Actinaria, Hydrozoa, Milleporina and Tridacna) from inshore, outer-reef and high latitude locations (Figure 1, additional collection sites are indicated by stars). Here we present the first stage of the project, Symbiodinium clade distribution of the GBR. Eventually we will add subclade information and the additionally obtained Symbiodinium identities from inshore and outer reefs from the Far Northern and Southern GBR.
With this information we aim to establish an index that provides estimates of relative sensitivity of reef invertebrate communities. The project will therefore generate important information for management authorities since it increases our measure of relative regional sensitivity of reef invertebrate communities as well as providing an assessment of community change related to climate driven stress.
Attachments
- Documentdatabase-references.doc (40 KB)