
 Mr David Welch (James Cook University), David Welch
Mr David Welch (James Cook University), David Welch
Description
Holothuria whitmaei
Introduction to black teatfish and why they are important
Holothuroids (all sea cucumbers) in general play important ecological roles (Purcell et al. 2016). One role is remineralising large quantities of organic nutrients through their feeding and burying activities (Uthicke, 2001a), which increases the benthic productivity of coral reefs (Uthicke & Klumpp 1998; Uthicke 2001b). Further, the role of sea cucumbers in the dissolution of CaCO3 sediment provides an important source of alkalinity and may play a role in buffering ocean acidification at least at local scales on coral reefs. (Schneider et al. 2011; Purcell et al. 2016).
Black teatfish are one of the highest value sea cucumber species and so fisheries have been very important in the past in Torres Strait and the Great Barrier Reef. They prefer coral reef habitat and are most dense on reef flats though can be found on back slopes and reef fronts. Due to overfishing black teatfish became a prohibited species on the Great Barrier Reef in 1999 and in the Torres Strait in 2003 (Anon, 2011; Uthicke et al, 2004).
Black teatfish are one of the few species that spawn during winter (Shiell and Uthicke, 2006). Although knowledge of the early life history of black teatfish is poor, egg development in holothurians is generally short (24 hrs) and the planktonic larval duration varies among species and for some of the key harvested species ranges from 12 - 22 days (Ramofafia et al, 2003). Growth rates of sea cucumbers are also poorly understood but based on a study of black teatfish are generally believed to be slow with low overall productivity (Uthicke et al, 2004). Aging of holothurians in the wild has not been possible however modelling by Uthicke et al (2004) suggested that H. whitmaei are long-lived (potentially several decades), with have low and sporadic recruitment. This was due to the apparent slow rate of population recovery after overfishing.
Key concepts that relate to connectivity
Population genetic techniques showed that populations of black teatfish on individual reefs in the Great Barrier Reef are highly connected and that even populations from West Australia and on reefs in the Coral Sea are potential sources of recruits (Uthicke & Benzie, 2000, 2003). This is not likely to reflect contemporary scales given the low mobility of adults and the relatively short larval duration. Recent assessments on the impacts of climate change on fisheries species determined that, being a winter spawner, the reproductive success of black teatfish may be compromised and any such impacts will be evident in more northern tropical regions first such as Torres Strait (Welch and Johnson, 2013).
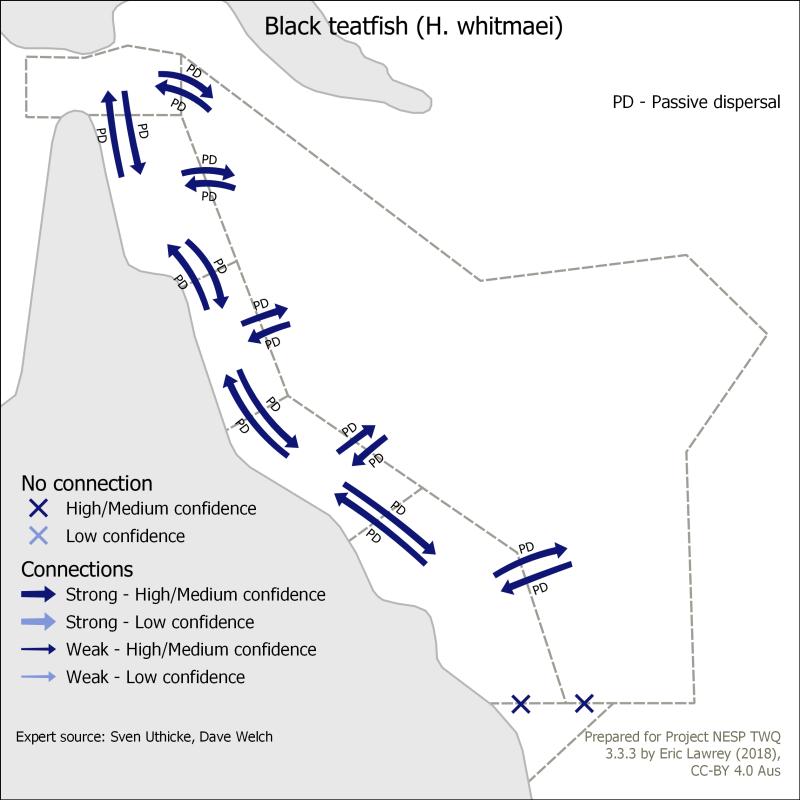 Dr Eric Lawrey (eAtlas (Australian Institute of Marine Science)), NESP TWQ 3.3.3, Sven Uthicke, Dave Welch
Dr Eric Lawrey (eAtlas (Australian Institute of Marine Science)), NESP TWQ 3.3.3, Sven Uthicke, Dave Welch
Figure 2. This map shows the distribution of black teatfish along the east coast and their connectivity among the different management jurisdictions. It was developed based on expert opinion by Karlo Hock.
References
- Anon (2011). Assessment of the Torres Strait Bêche-de-mer Fishery. Department of Sustainability, Environment, Water, Population and Communities report, Canberra, ACT, Australia, June 2011.
- Purcell, S.W., Conand, C., Uthicke, S. and Byrne, M. (2016) Ecological roles of exploited sea cucumbers. Oceanography and Marine Biology: An Annual Review, 2016, 54, 367-386
- Ramofafia, C., Byrne, M. And Battaglene, S.C. (2003) Development of three commercial sea cucumbers, Holothuria scabra, H. fuscogilva and Actinopyga mauritiana: larval structure and growth. Marine and Freshwater Research, 54(5): 657-667.
- Schneider, K., Silverman, J., Woolsey, E., Eriksson, H., Byrne, M. and Caldeira, K. (2011) Potential influence of sea cucumbers on coral reef CaCO3 budget: A case study at One Tree Reef. Journal of Geophysical Research 116: G04032, doi:10.1029/2011JG001755.
- Shiell, G.R. and Uthicke, S. (2006) Reproduction of the commercial sea cucumber Holothuria whitmaei [Holothuroidea: Aspidochirotida] in the Indian and Pacific Ocean regions of Australia. Marine Biology, 148(5): 973-986.
- Uthicke, S. (2001a) Nutrient regeneration by abundant coral reef holothurians. Journal of Experimental Marine Biology and Ecology, 265: 153–170.
- Uthicke, S. (2001b) Interactions between sediment-feeders and microalgae on coral reefs: grazing losses versus production enhancement. Marine Ecology Progress Series, 210: 125–138.
- Uthicke, S. and Benzie, J.A.H. (2000) Allozyme electrophoresis indicates high gene flow between populations of Holothura (Microthele) nobilis (Holothuroidea: Aspidochirotida) on the Great Barrier Reef. Marine Biology 137: 819-825.
- Uthicke, S. and Benzie, J.A.H. (2003) Gene flow and population history in high dispersal marine invertebrates: mitochondrial DNA analysis of Holothuria nobilis (Echinodermata: Holothuroidea) populations from the Indo-Pacific. Molecular Ecology 12: 2635-2648.
- Uthicke, S. and D.W. Klumpp. (1998) Microbenthos community production in sediments of a near shore coral reef: seasonal variation and response to ammonium recycled by holothurians. Marine Ecology Progress Series, 169: 1–11.
- Uthicke, S., Welch, D.J. and Benzie, J.A.H. (2004). Slow growth and lack of recovery in overfished holothurians on the Great Barrier Reef: Evidence from DNA fingerprints and repeated large-scale surveys. Conservation Biology, 18(5): 1395-1404.
- Welch, D.J. and Johnson, J.E. (2013) Assessing the vulnerability of Torres Strait fisheries and supporting habitats to climate change. Report to the Australian Fisheries Management Authority. C2O Fisheries, Australia (114pp.).